Genetic analysis recently identified nine new DNA repair genes of Aspergillus nidulans, musK - musS, which map on several different chromosomes (Käfer 1994, preceding article in this Newsletter). Such mus mutants are sensitive to certain chemical mutagens, but not sensitive or only slightly sensitive to UV and gamma-radiation. To identify epistatic interactions with members of the 4 Uvs groups, double mutants strains were isolated (Käfer and Chae 1994 Curr. Genet. 25:223-232). However, some mus;uvs double mutants could not be analyzed because they grew too poorly or were lethal (showing "synthetic enhancement in gene interaction"; Guarente 1993 Trends Genet. 9:362-366). The opposite effect was also found; namely interaction which led to improved recovery and growth (or "rescue") as documented here.
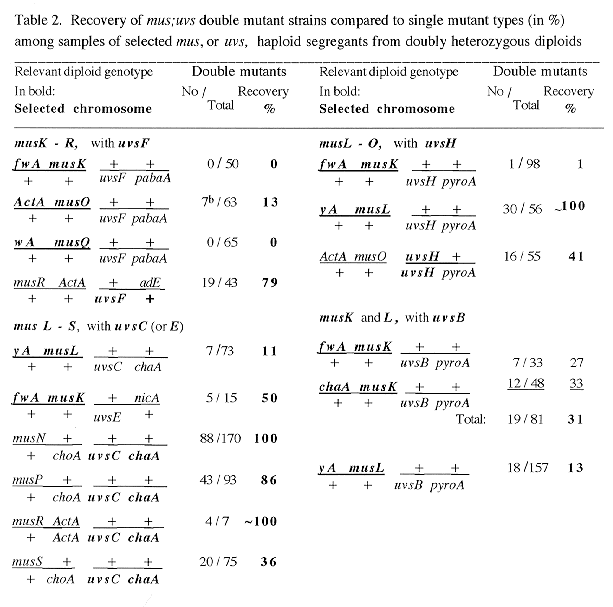
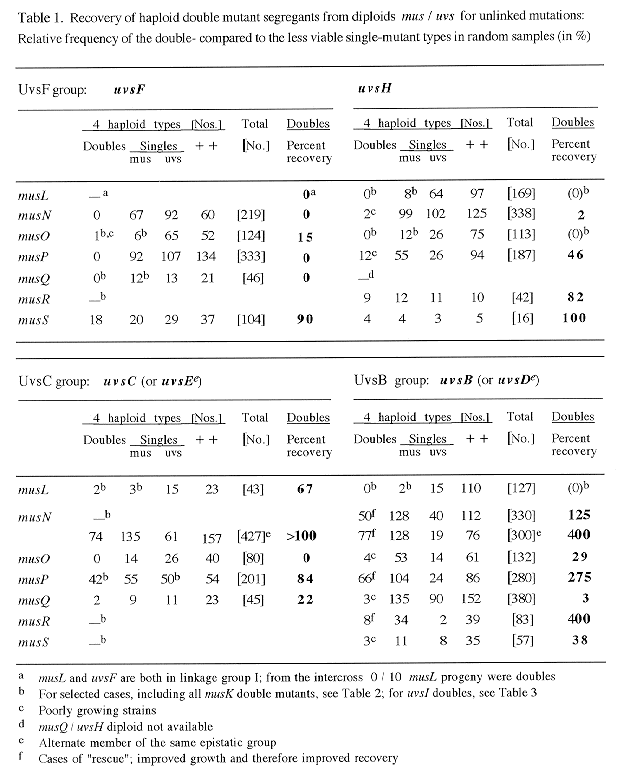
Double mus;uvs mutants were isolated as haploid segregants from heterozygous diploids when genes mapped on different chromosomes, or by random ascospore analysis of intercrosses in cases of linkage. Heterozygous diploids usually carried markers linked in repulsion to mus and uvs mutations. Samples of haploid segregants (50 to >300) were tested for nutritional markers and for growth on MMS, to which all mutants except uvsI are sensitive. In most cases, these tests identified the 3 mutant types, including the double mutant segregants which showed increased mutagen sensitivities in several cases (25% each ; Table 1).
In some cases recovery was poor even for single mutants (especially for musL or uvsB segregants). To obtain sufficiently large samples of poorly viable segregants, linked color markers or associated phenotypes were used to "select" the missing types (e.g., segregants were isolated preferentially for musL, musQ, musK and uvsC, which are linked to yA, wA, or to fwA and chaA, respectively; similarly, uvsB and D, or musO, could be recognized because they show dark mycelial color, or poor conidiation). In such selected samples, only single and double mutant segregants are expected (50% each; Table 2).
When marker segregation suggested that double-mutant segregants were much rarer than expected, additional haploids from at least two diploids were analyzed (one case with an aberration as the cause of low recovery was identified in this way; musQ;uvsH, missing in Table 4). In some cases, very rare segregants with markers expected of double mutants were crossovers, especially when recovery of single mutants was normal. Such exceptions, when checked on benomyl, occasionally turned out to be diploid (e.g., musO/+;uvsH/H, which was useful for the isolation of musO;uvsH double mutants; Table 2). In all cases, rare presumptive doubles and cases with ambiguous phenotypes were tested by outcrossing or by complementation to confirm the double mutant genotype.
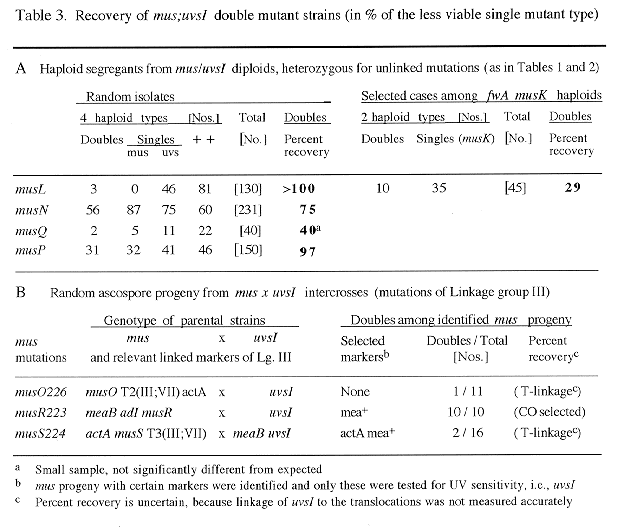
For some pairs of mus and uvs mutations which showed meiotic linkage, double mutant crossover types were obtained by selecting against linked markers used in repulsion (especially in crosses of the III;VII translocations associated with musO or S to uvsI which maps on chromosome III; Table 3). Double mus;uvsI mutants from such crosses were identified as MMS-sensitive progeny which also showed the high UV sensitivity typical for uvsI (Chae and Käfer 1993 Curr. Genet. 24:67-74).
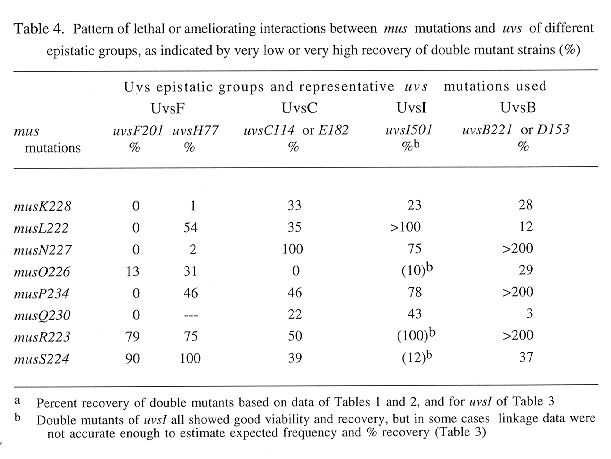
All types of isolated segregants are listed in Tables 1-3, and the recovery of double mutants is shown in percent relative to the "less viable" single mutant type. In general, only double mutants with good growth showed close to 100% recovery (e.g. mus;uvsI doubles), because of the competition between different segregant types on the benomyl plates during haploidization. "Lethal" interactions are assumed when practically no double mutants but significant numbers of both single mutant parents were obtained (e.g., in Table 1, for uvsF when combined with musN, or in Tables 1 and 2, for uvsF;musQ doubles). In random samples, certain musL and musO double mutants also could not be recovered, mainly because single mutants showed very poor recovery. However, when single mutants were selected, a significant fraction of doubles were obtained among them (compare results in Table 1 with those in Table 2).
The relative recoveries of double mutants from all the data are combined in Table 4. Clearly the majority of lethal interactions were found for mus mutations interacting with uvsF (5 of the 6 observed cases); only one other case is completely lethal (musO with uvsC). Unexpectedly, none of the mus;uvsH77 double mutants was completely lethal and only two showed very poor recovery, even though uvsH is a member of the UvsF epistatic group. In contrast, the two members of the UvsB group showed very similar results. Both uvsB and D affected viability of double mutants in all cases, but in two opposite ways for different mus mutations. Double mutants either were growing more poorly than uvsB (or D) singles and were produced with very low frequencies; or they showed better growth than uvsB (or D) and much increased recovery ( >200 - 400 % for musN, P and R double mutants).
Lethal interactions between uvs mutations had been found previously appeared to be the rule for mutations of different epistatic groups in Aspergillus (Käfer and Mayor 1986 Mutat. Res.161:119-134). However, viable double mutants of uvsI with other uvs mutations have been identified recently, even for intergroup pairs (Chae and Käfer 1993 Curr. Genet 24:67-74) as also found here for the many cases of mus;uvsI doubles. The latter results resemble the findings for double rad mutants of budding yeast, which usually are viable in all combinations. Only a few "synthetic lethals" have been found for radiation-sensitive mutants of this species; e.g., for certain rad52 mutations, when combined with unusual alleles of the excision repair gene RAD3, or of the topoisomerase gene TOP1 (Montelone et al. 1988 Genetics 119:289-301; Levin et al. 1993 Genetics 133:799-814). However, for many other important mutants, interacting mutations which lead to enhanced effects and lethality have recently been reported, and selective systems have even been devised (Bender and Pringle 1991 Mol. Cell. Biol. 11:1295-1305).
An interesting finding of the opposite type is the interaction of certain mus mutations with UvsB group mutations; namely, in double mutants with musN, P and R, suppression of the low growth rate and poor viability typical for UvsB group mutants was found. Such double mutants were also recovered more frequently (3-8x) and showed lower MMS sensitivity than single uvsB (or D) segregants (but only at low concentrations; Käfer and Chae 1994, ref. cit.). Furthermore, the same effect was observed in triple mutant strains. Growth rate and recovery improved (2-4 x) when musN was incorporated into double uvs strains containing a UvsB group mutation (e.g., from one such diploid, 18 musN;uvsC;uvsD, plus 66 musN;uvsD segregants were recovered, compared to 1 uvsC;D plus 19 uvsD not containing musN). In addition, triple mutant musN;uvsC;uvsD strains conidiate and grow much better than uvsC;uvsD double mutants which have extremely poor growth and conidiation. These findings are analogous to those known for certain recA mutants of E. coli in which the recBCD enzyme recklessly degrades DNA, while recA;recB double mutants do not show such effects.
Acknowledgement: This work was supported by a grant from the Natural Sciences and Engineering Research Council of Canada.