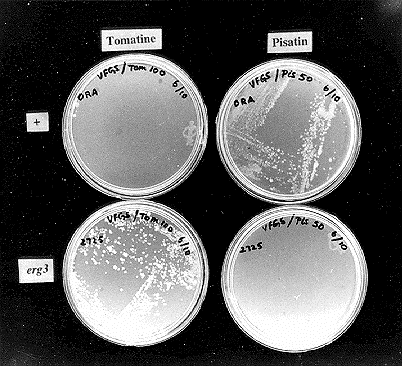
Figure 1: The phenotype of wild-type Neurospora crassa 74-OR23-IA (FGSC 987) and the erg-3 mutant (FGSC 2725) on Vogel's FCS medium supplemented with tomatine (100 g/ml) or pisatin (50 g/ml).
The erg-3 gene of Neurospora crassa encodes the sterol biosynthetic enzyme sterol 14,15 reductase. This was first suggested by the finding that the erg-3 mutant strain accumulated precursors of ergosterol that retain the double bond between carbons 14 and 15 in the sterol C ring (Ellis et al. 1991, J. Gen. Microbiol. 137:2627-2630). Consistent with this result, cloning and sequencing of erg-3 revealed that the encoded protein is highly homologous to the sterol 14, 15 reductase of yeast (Papavinasasundaram and Kasbekar, 1994, J. Genet. 73:33-41). The cloning of erg-3 (and of erg-1, the structural gene encoding sterol 8,7 isomerase) was made possible by the discovery that their mutants are sensitive to the pea phytoalexin pisatin (Papavinasasundaram and Kasbekar, 1993, J. Gen. Microbiol. 139:3035-3041).
Although the erg mutants were originally isolated by selection for resistance to the polyene antibiotic nystatin, the erg-3 mutant was only slightly (~2x) more nystatin-resistant than the wild-type. We report here that erg-3 mutants are also resistant to the tomato saponin alpha-tomatine (not to be confused with Tomatin the single malt scotch whisky) and this is a more reliable and convenient phenotype for scoring erg-3 mutations. Our choice of tomatine was prompted by a report that a mutation-induced 20% reduction in ergosterol content was sufficient to confer tomatine-resistance to the phytopathogenic fungus Fusarium solani (Defago and Kem, 1983, Physiol. Plant Path. 22:29-37). Interestingly, the mutant had now acquired the ability to infect green tomato fruits (which contain tomatine) whereas wild-type F. solani is nonpathogenic on tomato.
Figure 1 compares the growth of the mutant and wild-type Neurospora strains on Vogel's-FGS medium supplemented either with 100 ug/ml of tomatine or 50 ug/ml of pisatin. Stock solutions of tomatine (Sigma, 25 mg/ml) and pisatin (extracted from germinated pea seeds following the procedure of Sweigard and VanEtten, 1987, Phytopathology, 77:771-775, 55 mg/ml) were made in dimethyl formamide and dimethyl sulfoxide, respectively. As can be seen in this figure the wild-type is pisatin-resistant and tomatine-sensitive whereas the mutant has the converse phenotype. To confirm that the tomatine-resistance phenotype was in fact due to the erg-3 mutation (and not due to some unknown mutation in the erg-3 strain), we tested it for complementation by the cloned erg-3 gene. Mutant protoplasts were co-transformed with the plasmid pKGP86 (which carries the erg-3+ allele on a 2.1 kb KpnI-PstI insert derived from the cosmid G18:Al0 of the Orbach/Sachs library) and the plasmid pMP6 bearing the bacterial hpt gene. Transformants were isolated using a double selection for resistance to hygromycin 200 ug/ml) and pisatin (20 ug/ml). Conidia derived from the transformants were now tomatine-sensitive (data not shown), thereby confirming that the tomatine-resistance phenotype was due to the erg-3 mutation.
We also used the tomatine-resistance phenotype to identify new RIP-induced erg-3 mutations. A 1.5 kb HindIII fragment that was internal to the 2.1 kb KpnI-PstI insert of pKGP86 was subcloned into the vector pCSN44 (which carries the bacterial hpt gene) to produce the plasmid pSS17. This construct was used to transform the wild-type strain ORA. Transformants were selected for hygromycin-resistance and mated to the strain FGSC 7253 that is marked on linkage group III by acr-2 and trp-1. Random ascospores were harvested from such crosses and used to initiate vegetative cultures. Conidia from each culture were streaked on Vogel's-FGS medium containing 100 ug/ml tomatine. From each cross between 3 and 10% of the ascospores yielded tomatine-resistant and pisatin-sensitive cultures. This switched phenotype was due to the induction of new erg-3 mutations by RIP because it too could be complemented by the cloned gene (data not shown). This experiment verified that the tomatine-resistant, pisatin-sensitive phenotype of erg-3 mutants is independent of the genetic background of the strain in which the mutation is induced.
erg-3 thus represents the first fungal gene in which mutations cause a switch in the sensitivity between two plant antibiotics; tomatine and pisatin. We have recently cloned the sterol 14, 15 reductase gene from the pea pathogenic fungus Nectria haematococca MPVI (the perfect stage of F. solani) (D.P. Kasbekar and H.D. VanEtten, unpublished). It is now possible to construct similar mutants in Nectria by targeted gene disruption. If these mutants also display a switch in sensitivity between pisatin and tomatine might they also switch hosts from peas to tomatoes?
We thank Robert Sandrock (University of Arizona, USA) for drawing our attention to tomatine. This work is supported by the CSIR and the Department of Biotechnology, Government of India.
Figure 1: The phenotype of wild-type Neurospora crassa 74-OR23-IA (FGSC 987) and the
erg-3 mutant (FGSC 2725) on Vogel's FCS medium supplemented with tomatine (100
g/ml) or pisatin (50 g/ml).